
La α-lactoalbúmina es la principal proteína del lactosuero en la leche materna, alcanzando una concentración de 2,44 g/L en la leche madura. Su principal función es la síntesis de lactosa a partir de glucosa y galactosa en la glándula mamaria, aunque posee además otros efectos beneficiosos sobre la salud del lactante debido a su elevada proporción de aminoácidos esenciales (triptófano y cisteína). Según diversos estudios parece influir positivamente en la absorción de hierro en el intestino del niño, y en experimentos in vitro, unida al ácido oleico (complejo HAMLET), es efectiva frente a tumores celulares como el papiloma humano. El complejo HAMLET también presenta un claro efecto antimicrobiano frente a Streptococcus pneumoniae, Haemophilus influenzae, cepas enteropatógenas de Escherichia coli y Salmonella thypimurium, sin embargo no se ha demostrado que durante la digestión de la leche materna se forme dicho complejo en el tracto digestivo del lactante. El desarrollo de fórmulas infantiles destinadas a la alimentación del niño durante el primer año de vida ha mejorado considerablemente en las últimas décadas intentando no sólo adecuar la concentración de nutrientes a los requerimientos del lactante, sino también adicionando compuestos bioactivos de diferente naturaleza, como la α-lactoalbúmina, con el objetivo de alcanzar los efectos funcionales que se producen en los niños alimentados con leche materna.
Palabras clave: α-lactoalbúmina, efectos beneficiosos, fórmulas de inicio.
α-lactalbumin is the main whey protein in human milk rising 2,44 g/L in mature milk. It has a key function in the synthesis of lactose from glucose and galactose in the mammary gland although this compound has also other beneficial effects on the infant health due to the high proportion of essential aminoacids (tryptophan and cysteine). It seems also to increase iron absorption in the digestive track, and in in vitro experiments, linked to oleic acid (HAMLET complex), has shown anticarcinogenic effects against cellular tumor such as human papilloma. In addition, this complex has been reported to exhibit antimicrobial properties against Streptococcus pneumoniae, Haemophilus influenzae, enteropathogenic strains of Escherichia coli and Salmonella thypimurium. However, the in vivo synthesis of HAMLET complex during milk digestion has not been proved yet. Infant formula have been improved considerably during the last decades not only adapting nutrient concentrations to infants requirements but also by the addition of new bioactive ingredients such as α-lactalbumin, to have the same functional effect as in breast fed babies.
Key words: α-lactalbumin, infant formula, beneficial effects.
Departamento de Tecnología de los Alimentos, Nutrición y Bromatología. Facultad de Veterinaria, Universidad de Murcia 30100. Murcia, España.
Instituto de Nutrición Infantil Hero España S.A. Alcantarilla, Murcia. España.
La Sociedad Europea de Gastroenterología, Hepatología y Nutrición Pediátrica y la Federación Internacional de Sociedades de Pediatría, Gastroenterología, Hepatología y Nutrición se reunieron en el año 2005 con expertos en el área de nutrición infantil para definir las necesidades nutricionales en la infancia. Los datos de la composición de la leche materna de mujeres sanas y bien nutridas proporcionan una guía para establecer la composición de fórmulas infantiles. Sin embargo estos valores no deben ser tomados en sentido estricto ya que la composición de la leche materna varía entre individuos y con la etapa de lactación, habiendo además considerables diferencias entre la biodisponibilidad y efecto metabólico de nutrientes propios de la leche humana y los similares añadidos a las fórmulas infantiles. Por lo tanto una composición adecuada para una fórmula infantil se conseguirá comparando los efectos fisiológicos (patrón de crecimiento), bioquímicos (marcadores plasmáticos) y funcionales (como la respuesta inmune) que producen dicha fórmula y los de la leche materna. Este grupo internacional de expertos concluyó que la fórmula infantil sólo debe contener ingredientes en las cantidades que sirvan para un propósito nutricional o que aporten un beneficio al lactante (1), lo cual queda igualmente establecido en los principios generales para el establecimiento de la composición esencial de los preparados para actantes en la Norma del Codex Alimentarius STAN 72-1981. Los últimos avances que se han realizado se dirigen a la reducción y adaptación del contenido proteico y de fósforo, y a la adición de taurina, carnitina, nucleótidos, ácidos grasos de cadena larga y oligosacáridos (2).
Una de las principales diferencias entre la leche de vaca y la leche humana es su concentración proteica. La leche humana aporta 14-16 g/L de proteínas durante los primeros días de lactación, 8-10 g de proteínas /L a los 3-4 meses y de 7 a 8 g/L a partir del sexto mes (3), mientras que la leche de vaca contiene 32 g/L de proteínas (4). Por esta razón los primeros intentos para adaptar la leche de vaca a la alimentación del lactante se centraron en diluirla (5) para reducir la cantidad de proteínas y minerales y así disminuir la carga renal de solutos, lo cual constituía un factor limitante en la alimentación del niño debido a la inmadurez de estos órganos (6). Posteriormente, los estudios se centraron en resolver el predominio de proteínas del lactosuero en la leche materna en relación a las fórmulas infantiles, mediante la adición de proteínas del lactosuero de origen bovino. En la mujer, la concentración de proteínas del lactosuero es muy elevada en los primeros días de la lactación mientras que la concentración de caseína es casi inapreciable. Con el avance de la lactación, aumenta la síntesis de caseína y el volumen de leche producido por la glándula mamaria, disminuyendo la concentración total de proteínas del lactosuero. Por lo tanto se sabe que no existe una relación fija de proteínas del lactosuero con respecto a la caseína en la leche humana sino que ésta varía a lo largo de la lactación adecuándose a las necesidades del niño. El cociente proteínas del lactosuero:caseína citado más frecuentemente en las referencias bibliográficas es de 60:40, y es una aproximación media, aunque hay que tener en cuenta que éste cambia desde 80:20 en las primeras etapas de lactación hasta 50:50 al final de la misma (3).
Otro factor a tener en cuenta en el diseño de fórmulas infantiles, es la diferente composición de las proteínas del lactosuero de la leche bovina en relación a la humana, incluso su composición aminoacídica. Los aminoácidos que son esenciales para el lactante se han establecido teniendo en cuenta el perfil aminoacídico de la leche materna. En este sentido, la concentración de triptófano y cisteína es la mitad en la leche de vaca que en la humana, cuando se expresa como porcentaje de proteína total (5). El triptófano es precursor de la serotonina y melatonina, neurotransmisores que regulan el apetito, el humor, la percepción del dolor y el patrón de sueño (5, 7). Por otro lado, la cisteína es un precursor del glutatión (sistema antioxidante) y de la taurina (8), aminoácido que juega un papel importante en el desarrollo del cerebro. Pero la suplementación de las fórmulas con triptófano o cisteína (como aminoácidos libres) no parece ser una manera efectiva de resolver estos problemas, ya que es posible que no sean utilizados de forma eficiente para la síntesis de proteínas porque son absorbidos antes que otros aminoácidos que constituyen la dieta. Además, la leche humana contiene sólo un 5% de aminoácidos libres, principalmente ácido glutámico, y taurina (9).
Por todo ello, es necesaria la investigación de métodos adecuados para que los aminoácidos aportados por las fórmulas infantiles proporcionen un patrón de aminoácidos en sangre lo más parecido posible al de los lactantes alimentados con leche materna, aunque este objetivo resulta difícil de conseguir a partir de las proteínas de la leche de vaca. Uno de los principales inconvenientes se produce debido a que la principal proteína del lactosuero bovino es la β-lactoglobulina (una proteína ausente en el lactosuero de la leche materna), siendo la α-lactoalbúmina la principal proteína del lactosuero de la leche humana (Figura 1). Una de las propuestas tecnológicas para mantener la calidad biológica de la proteína, es asegurar un buen aporte de triptófano y reducir el contenido de proteínas mediante el enriquecimiento de la fórmula con alguna proteína rica en dicho aminoácido, como la α-lactoalbúmina (3).
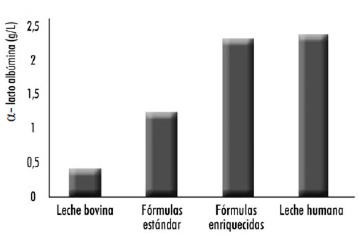
La proteína α-lactoalbúmina se encuentra en la leche de vaca en una concentración de 1 a 1,5 g/L, siendo aproximadamente el 3,4% de las proteínas totales y el 20% de las proteínas del lactosuero (10, 11). En la leche humana es la principal proteína del lactosuero, aumentando sus niveles desde un 21% hasta un 34% del día 1 al 14 de la lactación, y alcanzando en la leche madura (después del día 30) una concentración de 2,44 g/L (11, 12).
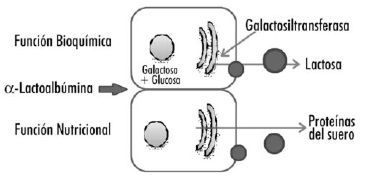
La α-lactoalbúmina posee una función bioquímica por su participación en reacciones enzimáticas, y una función nutricional ya que forma parte de la composición de la leche materna (Figura 2). La función bioquímica de esta proteína en las células de la glándula mamaria es de vital importancia en la lactogénesis, ya que junto a la galactosil-transferasa forma el complejo enzimático lactosa-sintasa, que cataliza la síntesis de lactosa a partir de glucosa y galactosa (13, 14). En concreto, la α-lactoalbúmina confiere a la galactosil-transferasa un incremento en la afinidad por la glucosa (15, 16), en contraposición a la formación de otros disacáridos que pueden ser sintetizados también por esta enzima. Una vez completada la síntesis de lactosa, la α-lactoalbúmina se disocia de la galactosil-transferasa (cuya unión es relativamente débil) y junto a otros componentes de la leche es transportado a la superficie apical de las células de la glándula mamaria, siendo descargada al lumen alveolar. La proteína es secretada a la leche siendo una importante fuente de aminoácidos para el lactante, cumpliendo su función nutricional (14). Su elevado contenido en aminoácidos esenciales (63% del total), particularmente triptófano (5,9% del total de los aminoácidos), cisteína y lisina, le confiere un importante valor nutricional para el lactante (17).
Las estructuras completas de la α-lactoalbúmina bovina y humana fueron inicialmente definidas por Brew et al., (18) y Findlay y Brew (19), respectivamente. Puesto que la función es semejante en ambas especies, sus estructuras son muy similares. Ambas están formadas por una cadena de 123 aminoácidos con un 72% de homología en la secuencia. Esta estructura tan parecida minimiza la posibilidad de que la α-lactoalbúmina bovina tenga propiedades antigénicas para el niño recién nacido si la comparamos con otras proteínas lácteas como la β-lactoglobulina.
La estructura nativa de la α-lactoalbúmina posee dos dominios; uno formado por una gran α hélice y otro por una pequeña hoja β, estos se unen por un puente de cisteína entre los residuos 73 y 91 que forman el sitio de unión del calcio. Otro puente disulfuro conecta los dos dominios en los residuos 61-77 y toda la estructura es estabilizada por cuatro puentes disulfuro (20). La α-lactoalbúmina posee un sitio de fuerte unión al calcio que influye directamente en su estabilidad, aunque no es esencial para su actividad dentro del complejo lactosa-sintasa. Posee además un sitio secundario de unión al mismo mineral al que se acoplan cuatro cationes. La unión al calcio produce cambios en la función y estructura terciaria de la proteína (21). Además de este mineral se pueden unir otros cationes como Mg2+, Mn2+, K+ y Na+, que compiten con el calcio produciendo en la misma cambios similares.
En las Tablas 1 y 2 se resumen algunos de los estudios (in vivo e in vitro, respectivamente) que avalan los efectos beneficiosos de la proteína α-lactoalbúmina y de los péptidos procedentes de su digestión, así como los relacionados directamente con la actividad funcional que puede aportar la proteína cuando se adiciona a alimentos infantiles. Los efectos sobre la salud que produce la α-lactoalbúmina pueden dividirse en tres grupos; aquellos relacionados con la proteína intacta, los producidos por los péptidos obtenidos tras la hidrólisis parcial de la proteína y por último los que producen los aminoácidos libres que resultan de la digestión completa de dicha proteína (11). La mayoría de las investigaciones se centran en este último punto debido a que la α-lactoalbúmina es particularmente rica en aminoácidos esenciales, sin embargo se debe destacar que existen otras funciones biológicas que derivan de la ingesta de α-lactoalbúmina.


Aunque, como ya se ha expuesto anteriormente, la estructura primaria de la α-lactoalbúmina bovina y humana es similar, se desconoce si se forman los mismos péptidos antimicrobianos durante la digestión in vivo. Además, es importante tener en cuenta que los procesos de purificación de esta proteína del lactosuero son cruciales para mantener su estructura y su bioactividad. El tratamiento térmico altera el patrón de uniones disulfuro entre proteínas y/o causa uniones cruzadas intermoleculares. La α-lactoalbúmina sola o en presencia de otras proteínas del lactosuero induce la formación de uniones intermoleculares por puentes disulfuro entre la proteína en sí, entre la α-lactoalbúmina y la β-lactoglobulina, o entre la α-lactoalbúmina y la albúmina sérica bovina, lo que impediría la liberación de los péptidos bioactivos con acción antimicrobiana.
El triptófano, es un aminoácido precursor de la serotonina cerebral y de la melatonina. Ambos compuestos regulan una gran cantidad de reacciones como el apetito, y la saciedad, la percepción del dolor, la depresión y el ritmo del sueño. En un estudio realizado en adultos, Markus et al., (22) compararon el efecto de la ingesta de α-lactoalbúmina frente a la caseína sobre la respuesta al estrés, comprobando que entre los individuos susceptibles al estrés, aquellos que habían tomado α-lactoalbúmina mostraban menos episodios de depresión que los que ingerían caseína, asociando este hecho a un aumento de la concentración de triptófano en plasma. En un estudio posterior Markus et al., (23) ratificaron el aumento de triptófano en plasma tras el consumo de una dieta rica en α-lactoalbúmina respecto a la dieta control, observándose mejor función cognitiva en pacientes vulnerables a padecer un estado de depresión. Por otro lado, los niños recién nacidos pueden encontrarse en riesgo de insuficiente biodisponibilidad de triptófano para la síntesis óptima de serotonina en el cerebro. En este sentido, en un estudio realizado en niños por Yogman et al., (7) aquellos que recibían triptófano entraban más rápidamente en episodios activos y de sueño que los que recibían valina o fórmula infantil estándar.
Los efectos beneficiosos de las fórmulas infantiles con concentraciones aumentadas en α-lactoalbúmina fueron mostrados por Heine et al., (24) en un estudio en el que una fórmula estándar, rica en proteína (18 g/L) y baja en α-lactoalbúmina fue comparada con otras dos fórmulas bajas en proteínas (13 g/L), y con una concentración intermedia y alta de α-lactoalbúmina. Sólo los recién nacidos que recibieron la fórmula con alta concentración de α-lactoalbúmina mostraron una concentración de triptófano sérico tan alto como los alimentados con leche humana. Este estudio demostró que el desarrollo de fórmulas infantiles bajas en proteínas pero enriquecidas con α-lactoalbúmina, supone un importante avance en el desarrollo de fórmulas infantiles más parecidas en composición a la leche humana. Sandström et al., (25) realizaron un estudio en recién nacidos sanos comparando un grupo alimentado con lactancia natural con otro alimentado con una fórmula estándar a base de proteínas del lactosuero y otras dos fórmulas ricas en α-lactoalbúmina (25%) con diferentes concentraciones de glicomacropéptidos (15% y 10%). Todas las fórmulas tenían la misma cantidad de proteína, sin embargo las fórmulas con 25% de α-lactoalbúmina contenían un 20% más de triptófano que la fórmula estándar. Este hecho dio lugar a una concentración plasmática del aminoácido similar al obtenido en los niños alimentados con leche materna. El nitrógeno ureico en sangre fue significativamente mayor en todos los niños alimentados con fórmula, en relación a los niños alimentados con leche materna y puesto que el crecimiento fue óptimo, estos autores propusieron disminuir la cantidad total de proteína en las fórmulas, siempre que ésta sea de alta calidad, ya que elevados niveles de proteína suponen una elevada carga renal. En este sentido Lien et al., (26) llevaron a cabo un estudio en el que evaluaron una fórmula con menor contenido proteico y enriquecida con α-lactoalbúmina, observando un patrón de crecimiento óptimo.
Son varios los estudios que investigan el efecto de las fórmulas infantiles enriquecidas con α-lactoalbúmina sobre el crecimiento de los lactantes y en todos los casos los resultados obtenidos demuestran un patrón de crecimiento más parecido al de los lactantes alimentados con leche materna que el de los niños alimentados con fórmulas estándar (25, 27, 28).
La unión de los minerales a péptidos procedentes de las proteínas del suero es menor y menos específica que la unión a péptidos originarios de la caseína, ya que tienen cargas negativas que unen eficientemente cationes divalentes (Fe2+, Mg2+, Mn2+, Cu2+ y Se2+). Sin embargo los péptidos que se producen mediante hidrólisis in vitro o in vivo de la proteína α-lactoalbúmina, captan los minerales actuando de transportadores pudiendo aumentar subiodisponibilidad (29). Así, en un estudio realizado en monos Rhesus, Kelleher et al., (30) observaron que las fórmulas enriquecidas con α-lactoalbúmina producían un aumento en la absorción de zinc con respecto a los animales alimentados con leche materna. También observaron un aumento en la absorción de hierro pero sin diferencias estadísticamente significativas respecto al grupo control. Sin embargo, Sändstrom et al., (25) comprobaron un incremento en la absorción de hierro en niños alimentados con fórmulas ricas en α-lactoalbúmina (25%). Este aumento se reflejó en los niveles de hierro sérico, sin aparecer modificaciones en la ferritina sérica ni en el hematocrito. La concentración de minerales en las fórmulas es superior a la de la leche materna debido en muchos casos a su reducida absorción, pero la adición de α-lactoalbúmina podría conseguir una mayor biodisponibilidad de cationes divalentes, siendo necesario añadir una menor proporción del mineral a las fórmulas infantiles.
En 1995 se observó una nueva actividad de esta proteína que despertó gran interés, Hakansson et al., (31) encontraron una forma multimérica de la α-lactoalbúmina con actividad antitumoral selectiva. Posteriormente se demostró que esta proteína unida al ácido graso C18:1:9 cis (ácido oleico) formaba un complejo (HAMLET/BAMLET: Human or Bovine Alpha-lactalbumin Made Letal to Tumor cells) que también poseía actividad in vitro frente a diferentes tipos de tumores (32) induciendo apoptosis celular. Además, recientemente se ha demostrado in vivo el efecto terapéutico específico del complejo HAMLET sobre varios tumores como glioblastomas humanos, tumores en glándulas mamarias de ratón y también sobre el papiloma humano en el que se observó una reducción de las lesiones causadas por dicho virus en un 100% de los pacientes tratados con el complejo HAMLET (33).
Sin embargo, hasta ahora los beneficios sobre la salud de los neonatos de los productos derivados de la digestión de la leche, y la posible formación los complejos HAMLET en alguna etapa determinada de la digestión, permanece sin esclarecer (11).
En dos recientes estudios in vitro (34, 35) se ha demostrado la actividad positiva de las proteínas del lactosuero sobre el sistema inmune. Por otro lado estudios con cultivos celulares y estudios in vivo han demostrado que estas proteínas son capaces de aumentar la respuesta inmune no específica y específica (36, 37). La alta concentración de aminoácidos precursores de glutatión parecer ser la causa de los efectos inmunológicos producidos (38).
Según diversos autores, la α-lactoalbúmina estimula el crecimiento de bifidobacterias, uno de los grupos microbianos indicadores de la salud intestinal del niño. Este hecho fue observado por Kee et al., (39) en un estudio in vitro realizado con Bifidobacterium longum ATCC 15707 y los péptidos resultantes de la digestión de la proteína. Además, estos péptidos poseen un efecto inhibitorio sobre Bacteroides, Clostridium y E. coli, llegando a ser la reducción de estas bacterias potencialmente patógenas semejante a la que se observa en recién nacidos alimentados mediante lactancia materna (40). En un estudio in vivo realizado por Bruck et al., (41) se observó un ligero efecto bifidogénico en la población de niños alimentados con fórmulas enriquecidas con α-lactoalbúmina sólo en aquellos casos en los que la población de bacterias beneficiosas era en inicio muy baja (niños no alimentados con leche materna).
Son diversos los estudios in vitro que demuestran la actividad antimicrobiana de los péptidos que se producen tras la digestión de esta proteína Pihlanto-Leppälä et al., (42) indicaron que tanto la pepsina como la tripsina liberan péptidos a partir de la α-lactoalbúmina capaces de inhibir el crecimiento de Escherichia coli, mientras que la misma proteína sin ser sometida a hidrólisis carece completamente de efecto. Se demostró el efecto antimicrobiano que ejerce la α-lactoalbúmina, cuando es añadida a fórmulas infantiles, frente a bacterias patógenas inoculadas en monos Rhesus (Escherichia coli enteropatógeno y Salmonella thypimurium) (43). Por otro lado, tres de los polipéptidos liberados tras la digestión (mediante la acción de la tripsina y la quimotripsina) presentaron efecto antimicrobiano frente a Klebsiella pneumoniae, Staphylococcus aureus, Staphylococcus epidermidis, Streptococci y C. albicans (44). Brück et al., (45) demostraron que las células Caco-2 expuestas a péptidos procedentes de la digestión tanto de la proteína α-lactoalbúmina como de glicomacropeptidos mostraban menor adhesión de los patógenos Eschechia coli enteopatógeno, Shigella flexneri y Salmonella thiphymurium respecto a las células control.
Entre otras funciones atribuidas a los péptidos procedentes de la digestión de esta proteína se encuentra la actividad reductora de la presión sanguínea. Diversos estudios demuestran que diferentes péptidos procedentes de la proteína α-lactoalbúmina sometida a digestión enzimática, poseen actividad antihipertensiva (46, 47). Se ha demostrado también la capacidad antioxidante de los péptidos obtenidos de la digestión de la α-lactoalbúmina (48), las propiedades antiulcerativas y su actividad opioide (49, 50).
El enriquecimiento de las fórmulas infantiles con α-lactoalbúmina permite obtener un producto con una composición proteica y aminoacídica más parecida a la leche humana, hecho que tiene una gran importancia desde un punto de vista nutricional al ser esta proteína una fuente natural de triptófano y cisteína. Además, es importante valorar la actividad biológica beneficiosa que puede producirse en el recién nacido alimentado con lactancia artificial con la incorporación de α-lactoalbúmina en su dieta, principalmente por su acción inmunoestimuladora y antimicrobiana, ya que en edades tempranas el sistema inmunológico no se encuentra suficientemente desarrollado, siendo mayor la incidencia de enfermedades gastrointestinales que los niños lactados a pecho.
Recibido: 13-12-2011
Aceptado: 13-04-2012